DNR transkripcija: DNR transkripcijos procesas ir mechanizmas
Perskaitykite šį straipsnį, kad sužinotumėte apie DNR transkripciją: DNR transkripcijos procesą ir mechanizmą
Genetiškos informacijos kopijavimas iš DNR antisenso arba šablono grandinės į RNR vadinamas transkripcija. Jis skirtas koduotai informacijai paimti iš DNR į vietą, kur ji reikalinga baltymų sintezei. Papildomumo principai naudojami net transkripcijos metu.
Išimtis yra ta, kad (i) Uracilas yra vietoj timino, esančio priešais šablono adeniną, (ii) Tik transkribuojama tik DNR šablonas. Abi DNR grandinės negali būti kopijuojamos transkripcijoje, nes tai sukurs dviejų tipų baltymus, po vieną su teisinga aminorūgščių seka ir kita su atvirkštine aminorūgščių seka.
Be to, jei tuo pačiu metu gaminami du papildomi RNR, jie turėtų tendenciją suformuoti dvigubos RNR, dėl kurios koduotoji informacija netaptų baltymų. Visas transkripcijos pratimas tuomet atrodytų nereikalingas.
Transkripcijos vienetas:
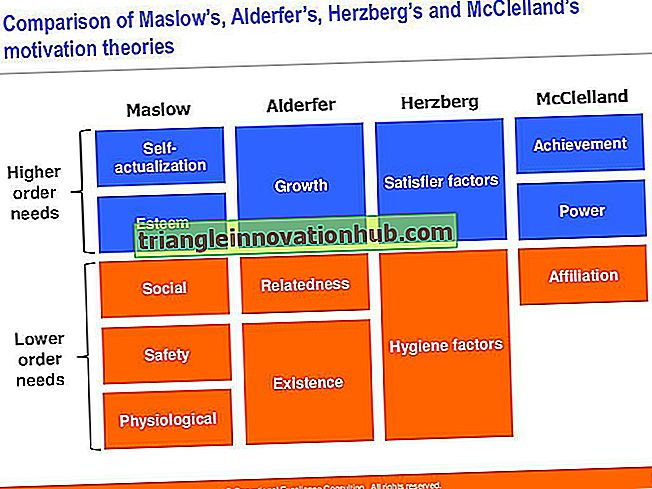
DNR segmentas, dalyvaujantis transkripcijoje, vadinamas transkripcijos vienetu (6.16 pav.). Jame yra trys komponentai (i) promotorius, (ii) struktūrinis genas ir (iii) terminatorius. Be promotoriaus, eukariotams reikalingas stiprintuvas. Reklamuotojas yra prieš struktūrinį geną. Pagal susitarimą jis vadinamas 5 'galu (koduojančios juostos, kuri yra 3 ′ šablono juostos galas). Terminatoriaus regionas yra pasroviui nuo struktūrinio geno 3 'gale (kodavimo grandinės, kuri iš tikrųjų yra 5' šablono juostos galas). Reklamuotojas turi skirtingas dalis, skirtas prijungti prie įvairių transkripcijos faktorių.
Daugeliu atvejų reklamuotojas turi AT turtingą regioną, vadinamą TATA. Srityje yra griovelis, į kurį galima sujungti konkrečius baltymų komponentus. TATA turintis regionas taip pat vadinamas Pribnow langeliu po jo atradėjo pavadinimo.
Struktūrinis genas yra tos DNR grandinės komponentas, kuris turi 3 '→ 5' poliškumą (kadangi transkripcija gali vykti tik 5 '→ 3' kryptimi). Ši DNR kryptis vadinama šablonine grandine arba pagrindine grandine arba antisense arba (-) grandine. Kitą grandinę, kurios poliškumas yra 5 '→ 3 ′, transkripcijos metu yra perkelta. Ši nesuskaitoma kryptis, kuri nedalyvauja transkripcijoje, taip pat vadinama jutimo arba kodavimo grandine arba pliusu (+), nes šioje eilutėje esantis genetinis kodas yra panašus į genetinį kodą (remiantis mRNR), išskyrus tai, kad uracilas yra pakeistas tyminu.
Transkripcijos mechanizmas:
Eukariotuose transkripcija vyksta per I fazę diferencijuotose ląstelėse, bet daugiau - G1 ir G2 fazėse ląstelių ciklo viduje branduolyje. Priklausomai nuo reikalavimo, struktūrinis genas gali perrašyti vieną į daugelį RNR molekulių. Transkripcijos produktai persikelia į citoplazmą.
Prokariotuose transkripcija vyksta kontaktuojant su citoplazma, nes jų DNR yra citoplazmoje. Transkripcijai reikia DNR priklausomos RNR polimerazės. Eukariotuose yra trys RNR polimerazės, Pol I (Pol A) (ribosolio arba rRNR, išskyrus 5S rRNR), Pol II (mRNR, snRNS) ir Pol III (perdavimo arba tRNR, 5S rRNR ir kai kurių snRNA). Eukariotinėms RNR polimerazėms pradėti reikalingi transkripcijos faktoriai.
Įvairios DNR dalys yra susijusios su įvairių ribonukleino rūgščių transkripcija. Prokariotai turi tik vieną RNR-polimerazę, kuri sintezuoja visų tipų RNR. Escherichia coli Rna polimerazėje yra penkios polipeptidinės grandinės β, β ', α, α' ir σ (sigma) faktoriaus. Holoenzimo molekulinė masė yra 4, 50 000. Sigma arba faktorius atpažįsta DNR pradžios signalą arba promotoriaus regioną (TATA).
Polimerazės fermento dalis, atėmus faktorių, vadinama pagrindiniu fermentu (6.17 pav.). α ir α'-polipeptidai yra apsauginiai, o β ir β 'yra kataliziniai.
Norint nutraukti transkripciją, reikalingas nutraukimo faktorius, vadinamas Rho (p). Taip pat reikalingi keletas kitų veiksnių - DNR dvipusio atplėšimo, nesuvyniotos DNR grandinės stabilizavimo, bazinės poravimo, atskyrimo ir transkribuotos RNR apdorojimo.
1. Ribo-nukleotidų aktyvinimas:
Ribonukleotidai skiriasi nuo deoksiribonukleotidų, turintys ribozės cukrų vietoj deoksiribozės cukraus. Tymidino monofosfatas pakeičiamas uridino monofosfatu. Keturios ribonukleotidų rūšys yra adenozino monofosfatas (AMP), guanozino monofosfatas (GMP), uridino monosfosfatas (UMP) ir citidino monofosfatas (CMP). Jie atsiranda laisvai nukleoplazmoje. Prieš transkripciją nukleotidai aktyvuojami fosforilinimo būdu. Fermentinė fosforilazė reikalinga kartu su energija. Aktyvuoti arba fosforilinti ribonukleotidai yra adenozino trifosfatas (ATP), guanozino trifosfatas (GTP), uridino trifosfatas (UTP) ir citidino trifosfatas (CTP).
2. DNR šablonas:
Konkrečiais signalais DNR segmentai, atitinkantys vieną ar kelis cistronus, tampa demresuoti ir pasiruošę perrašyti. Kiekviename tokiame DNR transkripcijos segmente yra promotoriaus regionas, pradžios vieta, kodavimo regionas ir terminatoriaus regionas. Transkripcija prasideda inicijavimo vietoje ir baigiasi terminatoriaus regione. Promotoriaus regione yra RNR polimerazės atpažinimo vieta ir RNR polimerazės surišimo vieta.
Grandinės atidarymas vyksta TATAATG nukleotidų (TATA langelis) užimtuose regionuose daugelyje prokariotų. Fermentai, reikalingi grandinės atskyrimui, yra nelindazės, girazės ir viengubos rišamosios baltymai. Terminatoriaus regione yra arba poli A bazės seka, arba palindrominė seka (identiška pagrindo seka, veikianti priešinga kryptimi dviejose DNR grandinėse).
RNR polimerazė (paplitusi procariotuose ir būdinga eukariotams) jungiasi prie promotoriaus regiono. Dvi DNR krypties palaipsniui išsiskiria nuo polimerazės surišimo vietos. Vienas iš dviejų DNR krypčių (3'— »5 ') veikia kaip šablonas RNR transkripcijai. Tai vadinama pagrindine, šablono ar antisenso grandine. Transkripcijos formavimas vyksta 5 '-> 3' kryptimi.
3. Pagrindų poravimas:
Ribonukleozido trifosfatai, esantys aplinkinėje terpėje, atsiduria priešinga DNR šablono azoto bazėms (antisense grandinei). Jie sudaro papildomas poras, U priešais A, A priešais T, С priešais G ir G priešais C. Pirofosfatas išsiskiria iš kiekvieno ribonukleozido trifosfato, kad susidarytų ribonukleotidas. Pirofosfatas hidrolizuojamas fermento pirofosfatazės pagalba. Jis išskiria energiją.
4. Grandinės formavimas:
Naudojant RNR polimerazę, gretimos ribo-nukleotidai, laikomi per DNR šabloną, prisijungia, kad suformuotų RNR grandinę. Prokariotuose vienas polimerazė atpažįsta promotorių ir iniciacijos regioną. Eukariotuose yra atskiri transkripcijos faktoriai ir RNR polimerazė transkripcijai aktyvuoti. RNS grandinės formavimosi pradžioje prokariotinės RNR polimerazės sigmos (a) faktorius atskiria. RNR polimerazė (šerdies fermentas) juda palei DNR šabloną, sukeldama RNR grandinės pailgėjimą maždaug 30 nukleotidų per sekundę greičiu. RNR sintezė sustoja, kai polimerazė pasiekia terminatoriaus regioną. Tam reikalingas Rho faktorius (p). Terminatoriaus regione yra sustabdymo signalas. Jis taip pat turi 4-8 A-nukleotidus.
5. RNR atskyrimas:
Nutraukimas arba rho faktorius turi ATP-ase aktyvumą (Roberts, 1976). Jis padeda išlaisvinti užbaigtą RNR grandinę. Išleistas RNR vadinamas pirminiu transkriptu. Jis apdorojamas funkcinėms RNR formuoti. Daugelyje prokariotų, kai kurie susijusių funkcijų struktūriniai genai yra grupuojami operonuose. Operonas transkribuojamas kaip vienas vienetas. Toks transkripcijos įrenginys sukuria policistroninę mRNR. Eukariotuose transkripcijos vienetas duoda monokistroninę mRNR.
6. Duplex formavimas. Išleidus pirminį transkriptą, dvi DNR kryptis sukuria ryšius tarp papildomų bazinių porų. Atsiranda girazės, nesukilusios ir SSB baltymai. Todėl dukart spiralinė DNR forma atnaujinama.
7. Po transkripcijos apdorojimas:
Pirminis transkriptas dažnai yra didesnis už funkcines RNR. Šis pirminis transkriptas yra vadinamas heterogenišku branduoliniu RNR arba hnRNR, ypač mRNR atveju. Po transkripcijos apdorojimas yra būtinas visų tipų RNR pirminio transkripto konversijai į funkcines RNR (6.18 pav.). Tai keturių tipų:
i) Skilimas:
Didesni RNR pirmtakai yra suskaldyti, kad susidarytų mažesnės RNR. Pirminis rRNR transkriptas yra 45 S eukariotuose. Jis yra suskirstytas taip, kad susidarytų:
Pirminį transkriptą taip pat skaldo rfoonuclease-P (RNR fermentas). Pirminis transkriptas gali sudaryti 5-7 tRNR prekursorius.
ii) suskirstymas:
Eukariotiniai transkriptai turi papildomų segmentų, vadinamų intronais arba tarpinėmis sekomis arba nekoduojančiomis sekomis. Jie nerodomi subrendusioje ar perdirbtoje RNR. Funkcinės kodavimo sekos vadinamos eksonais. Sujungimas yra intronų pašalinimas ir eksonų suliejimas, siekiant sudaryti funkcines RNR. Kiekvienas intronas prasideda nuo dinukleotido GU ir baigiasi dinukleotido AG (GU-AG taisyklė).
Juos atpažįsta „Sn-RNPs“ (išreikštas kaip snurps) arba smulkių branduolinių ribonukleoproteinų (vizualizuoja Ul, U2, U4, U5, U6) jungiamųjų aparatų komponentai. Tarp introno 5 ′ galo (GU) ir 3 ′ galo (AG) yra suformuotas kompleksas, vadinamas spliceosome. Energija gaunama iš ATR. Ji pašalina introną. Gretimi egzonai yra sujungti. Galai uždaromi RNR ligaze (6.18 pav.).
Intronai nėra naujausia raida. Jie atsirado, kai buvo įdiegta RNR centrinė genetinė technika. Todėl suskaidyti genai ir skaidyti transkriptai yra senovės genetinės sistemos savybės. Skirstymas ir toliau yra RNR katalizinė funkcija. Daugelis tokių RNR priklausomų procesų pasirodo.
(iii) Terminalo papildymai (viršutinė riba ir uodegos):
Papildomi nukleotidai pridedami prie RNR galų specifinėms funkcijoms, pvz., CCA segmentas tRNR, kapsulės nukleotidai 5 m gale mRNR arba poli-A segmentai (200-300 liekanų) 3R gale. Dangtelis formuojamas modifikuojant GTP į 7-metil-guanoziną arba 7 mG.
(iv) Nukleotidų modifikacijos:
Jie dažniausiai pasitaiko tRNR - metilinimo (pvz., Metilcitozino, metil guanozino), deaminacijos (pvz., Inozino iš adenino), dihidrouracilo, pseudouracilo ir kt.
Prokariotuose mRNR nereikalauja, kad jis taptų aktyvus. Be to, transkripcija ir vertimas vyksta tame pačiame regione. Tai sukelia vertimo pradžią net prieš visiškai suformavus mRNR.
RNR in vitro sintezę pirmą kartą atliko Ochoa (1967).