DNR replikacija: DNR replikacijos mechanizmai
Kai kurie svarbiausi DNR replikacijos būdai yra tokie!
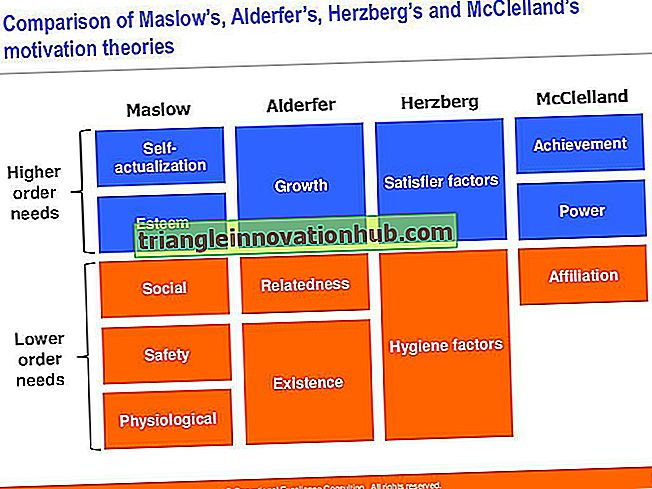
DNR replikacija eukariotuose yra pusiau konservatyvus, pusiau nepertraukiamas ir dvikryptis, lyginant su semikonservatyviu, dvikrypčiu ir nepertraukiamu prokariotuose.

Image Courtesy: dbriers.com/tutorials/wp-content/uploads/2012/12/DNA_replication_split.png
DNR replikacija vyksta ląstelių ciklo S fazėje. Tai daugiapakopis kompleksinis procesas, kuriam reikia daugiau kaip dešimties fermentų ir baltymų faktorių. Jis prasideda konkrečioje vietoje, vadinamoje replikacijos pradžia arba ori. Bakterinė ir virusinė DNR turi vieną replikacijos pradžią. Jis veikia kaip vienas atkartojantis vienetas arba replikonas.
Eukariotinėje DNR yra daug replikacijos pradžios. Jis turi kelis atkartojamus segmentus arba replikonus, ty daugiašakius. Nesant ori, replikacija nebus. Rekombinantinės DNR technologijos vektoriaus reikalavimas yra gauti replikacijos pradžią.
DNR replikacija yra labai brangi. Pagrindinis DNR replikacijos fermentas yra DNR priklausoma DNR polimerazė. DNR replikacija yra gana greita. E. coli DNR replikacija su 4, 6 x 106 bp reikalauja 19 minučių.
Vidutinė bazių polimerizacijos norma yra 2000 bp per sekundę kiekvienoje kryptimi. Replikacijai reikia gausios energijos, kuri atsiranda dezoksiribonukleotidų trifosfatų skaidymo metu.
Replikacija vyksta taip:
1. Deoksiribonukleotidų aktyvinimas:
Dezoksiribonukleotidai arba deoksiribonukleozido monofosfatai laisvai atsiranda nukleoplazmoje. Jie yra keturių tipų - deAMP (deoksiadenozino monofosfatas), deGMP (deoksiguanozino monofosfatas), deCMP (deoksicitidino monofosfatas) ir deTMP (deoksitimidino monofosfatas). Pirmiausia jie fosforiliuoti ir pakeisti į aktyvias formas, kuriose yra trys fosfato liekanos, o ne viena. Fermentų fosforilė yra reikalinga kartu su energija.
Fosforilinti nukleotidai yra deATP (deoksiadenozino trifosfatas), deGTP (deoksiguanozino trifosfatas), deCTP (deoksicitidino trifosfatas) ir deTTP (deoksitimidino trifosfatas). Šie bazių trifosfatai tarnauja dvigubai. Jie veikia kaip substratas, taip pat suteikia energijos nukleotidų polimerizacijai.

2. DNR grandinių ekspozicija:
Fermentinė helikazė (neindazė) veikia per Ori vietą ir išskleidžia dvi DNR sruogas, naikindama vandenilio jungtis. Atskiros juostos yra stabilizuojamos vienšūšiais rišamųjų baltymų (SS BP) arba spiralės stabilizuojančių baltymų pagalba. Atsipalaidavimas sukelia įtempimą nesupjaustytoje dalyje formuojant daugiau superkūšlių. Įtampa išsiskiria fermentų topoizomerazėmis.
Jie sukelia DNR grandinės nikuojamumą ir uždarymą. Kartu su topoizomeraze bakterijos turi kitą fermentą, vadinamą DNR giraze, kuri gali įvesti neigiamus superkūnius (vyresnio amžiaus darbuotojai manė, kad girazė veikia tiek helikazei, tiek topoizomerazei).
Įvairių fermentų pagalba abi DNR kryptis tampa atviros replikacijai. Tačiau dėl labai didelės energijos poreikio viena DNR neatidaro vienoje stadijoje. Atskyrimo taškas lėtai eina į abi puses. Kiekviena kryptimi ji suteikia Y formos struktūrą, vadinamą replikacijos šakele (6.13 ir 6.14 pav.).

3. RNR gruntas:
Tai labai svarbu pradedant naujas DNR grandines. RNR pradmuo yra nedidelė RNR kryptis, kuri yra sintezuojama naujos DNR grandinės 5 'gale, naudojant DNR specifinę RNR polimerazės fermentą, vadinamą primase. RNR gruntas yra suformuotas ant kitos krypties vienos grandinės ir šakės galo laisvo galo. RNR pradmenų susidarymas sudaro DNR sintezės pradžios fazę, nes be RNR pradmenų, DNR polimerazės negali pridėti nukleotidų.
Fagas ф x 174 ir kai kurios kitos prokariotinės sistemos reikalauja sudėtingesnio fermento, vadinamo primo. Eukariotuose primasės funkciją atlieka fermento DNR polimerazė α. Jis sukaupia ~ 10 bazinių RNR ir 20-30 DNR bazių (Lewin, 2004). Pradėjus nukleotidų grandinę, RNR pradmuo pašalinamas ir tarpas pripildomas DNR polimeraze I prokariotuose ir DNR polimerazėje β eukariotuose.
4. DNR polimerai:
Prokariotai turi tris pagrindinius DNR sintezės fermentų tipus, vadinamus DNR polimerazėmis III, II ir I. Visi jie prideda nukleotidų 5 '-> 3' kryptimi 3 ′ -> 5 ′ pradinės grandinės ruože. Jie taip pat turi 3 '-> 5 ′ ekso-nukleazės aktyvumą. Nors DNR polimerazė III yra daugiausia susijusi su DNR replikacija (naujų bazių pridėjimas ir polimerizacija), I polimerazė yra pagrindinis remonto fermentas. II polimerazė yra nedidelis remonto fermentas.
DNR polimerazė I taip pat turi 5 -> 3 eksonukleazės aktyvumą. Eukariotuose randamos penkių tipų DNR polimerazės - α, β, γ, δ ir ε, bet pagrindinės trys yra α, δ ir e. 8 polimerazė yra susijusi su pirmaujančios grandinės replikacija. Polimerazė gali padėti atsiliekančios grandinės sintezei kartu su kitais vaidmenimis. Polimerazė α yra didžiausias ir pagrindinis DNR replikacijos fermentas. Visos DNR polimerazės vienoje pusėje turi sukibimo rankos su nykščiu konfigūraciją, o pirštai - ant kitos ir palmių kaip įgaubta katalizinė vieta šablonams ir bazinėms poroms sujungti.
5. Suporavimas pagrindu:
Dvi atskirtos DNR grandinės replikacijos šakėje veikia kaip šablonai. Dezoksiribonukleozido trifosfatai atsiduria priešais veikiamų DNR šablonų azoto bazes - deTTP priešais A, deCTP priešais G, deATP priešais T ir deGTP priešais C.
Nukleofinė ataka atskiria pirofosfatą (PPi) nuo trifosfato. Nustatyti fosfodiesterio ryšiai. Pirofosfato hidrolizė fermentu pirofosfataze išskiria energiją. Dezoksiribilleozido trifosfatas → Dezoksiribilleozido monofosfatas + PPi

Energija naudojama nustatant vandenilio jungtis tarp laisvų nukleotidų ir šablonų azoto bazių.
6. Grandinės formavimas:
Jis reikalauja DNR polimerazės III (Kornberg, 1956) prokariotuose ir polimerazėje δ / ε eukariotuose. DNR polimerazė III yra kompleksinis fermentas, turintis septynis subvienetus (a, β, ƍ, ƴ, €, θ, τ). Esant Mg2 +, ATP (GTP), TPP ir DNR polimerazei III, gretimos nukleotidai, surasti prie kiekvieno šablono DNR grandinės azoto bazių, sukuria fosfodiesterio jungtis ir susiejasi su replikuota DNR grandine.
Kai replikacija vyksta, naujos pagrindinės DNR dvipusės sritys atsipalaiduoja ir atskiriamos, kad replikacija sparčiai vyktų iš kilmės vietos į kitą galą. DNR polimerazė I pašalina RNR pradmenį ir pripildo komplementarių nukleotidų spragą. Dėl DNR dvigubos grandinės nuoseklaus atidarymo ir jo replikacijos, sudarydama dvi grandines, DNR replikacija taip pat vadinama užtrauktuku.
Tačiau DNR polimerazė gali nukleotidus polimerizuoti tik 5 '→ 3' kryptimi 3 '-> 5' kryptyje, nes ji prideda juos 3 'gale. Kadangi dvi DNR kryptys, veikiančios priešparalelinėmis kryptimis, du šablonai turi skirtingus galus replikacijai. Tokiu būdu replikacija per du šablonus vyksta priešinga kryptimi. Viena kryptis su poliškumu У -> 5 ′ nuolat papildo grandinę, nes 3 'galas visada yra atviras pailgėjimui.
Tai vadinama pagrindine kryptis. Replikacija ant kito šablono su 5 ′ → 3 poliškumu yra nepertraukiama, nes tik mažas DNR krypties segmentas gali būti pastatytas 5 '→ 3 kryptimi dėl nedidelio šablono padėties vienu metu. Trumpi replikuotos DNR segmentai vadinami Okazaki fragmentais (= Okasaki segmentai; Reiji Okazaki, 1968). Kiekvienas iš jų turi 1000–2000 bp prokariotuose ir 100-200 bp eukariotuose.
RNR gruntas taip pat reikalingas kiekvieną kartą, kai bus pastatytas naujas Okazaki fragmentas. Pakeitus RNR pradmenį su dezoksiribonukleotidais ir jų polimerizaciją, Okazaki fragmentai sujungiami fermentu, DNR ligaze (Khorana, 1967). „Okazaki“ fragmentams sukaupta DNR kryptis vadinama atsilieka.
Kadangi viena kryptis nuolat auga, o kita kryptis susidaro nepertraukiamai, DNR replikacija yra pusiau nutraukta. Kadangi replikacija vyksta dviem kryptimis nuo replikacijos pradžios arba ori, viena iš tėvų krypčių sudarys pirmaujančią grandinę iš vienos pusės ir atsiliekančią juostą kitoje pusėje. Atvirkščiai yra antrojoje pusėje esančiose pagrindinėse dalyse. Tai padeda užbaigti replikaciją vienu metu visame replikone.
7. Patvirtinimas ir DNR taisymas:
Replikacijos metu kartais įvedama neteisinga bazė. Dažnis yra vienas iš dešimties tūkstančių. DNR polimerazė III gali suvokti tą patį. Jis grįžta atgal, pašalina neteisingą bazę, leidžia pridėti tinkamą bazę ir tada eina į priekį. Tačiau net ir DNR polimerazė III nesugeba atskirti uracilo nuo timino, todėl ji dažnai yra vietoj timino. Toks neatitikimas koreguojamas daugeliu fermentų.
Yra atskiras remonto mechanizmas dėl bet kokios DNR sukeltos žalos, atsirandančios dėl mutacijos, UV poveikio arba nesuderinamumo, kuris išsiskiria ištaisymo mechanizmu. Niksą ar pertrauką sukelia endonukleazė šalia remonto regiono. DNR polimerazė I (Komberg, 1969) pašalina nesuderintus arba neteisingus nukleotidus, jei jie yra, ir sintezuoja teisingą pakeitimą, naudodami nepažeistą grandinę kaip šabloną. Naujai suformuotas segmentas yra užsandarintas DNR ligaze.